Potassium homeostasis during exercise in domestic species: the role of the
sodium-potassium pump in skeletal muscle
Maria E. Everts
Department of Anatomy and Physiology, Faculty of Veterinary Medicine, Utrecht
University, PO Box 80.157, 3508 TD Utrecht, The Netherlands
Introduction
In 1997, Jens Christian Skou was awarded the Nobel Prize in Chemistry for
his discovery and elegant description, some 40 years earlier, of the
sodium-potassium (Na+,K+) pump in crab nerve fibres
[37]. It is now widely accepted that this cation transport system is essential
for cell function, and that it plays a central role in the
Na+,K+ homeostasis of virtually all animals [4],[6].
Since its identification, the Na+,K+ pump has been
the subject of numerous investigations, including ones on the mechanism for
controlling ion transport through the pump, the detailed molecular structure
of the pump and its regulation. Regulation of plasma K+, both
long-term (by kidney Na+,K+-ATPase), and acute (by
skeletal muscle Na+,K+-ATPase, during exercise), have
been questions frequently addressed by physiologists. Most of this work has
been performed using small rodents and man [4,5,6], however, the following
review will discuss the up- and down regulation of the
Na+,K+ pump concentration in the skeletal muscle of
domestic animal species, including cats, dogs, horses and cattle.
The Na+,K+ pump in skeletal muscle
To maintain the high concentration of K+ and the low concentration
of Na+ that exists in animal cells, relative to the external
environment, a specific transport system is required. Skou's discovery of
an enzyme that hydrolyses ATP only when Na+ and K+
are present (in addition to Mg2+ required by all ATP enzymes),
was the beginning of our understanding of the Na+K+
pump [37]. The enzyme, known as Na+,K+-ATPase, forms
an integral part of the Na+,K+ pump and the splitting
of ATP provides the energy required to drive the active transport of the
cations.
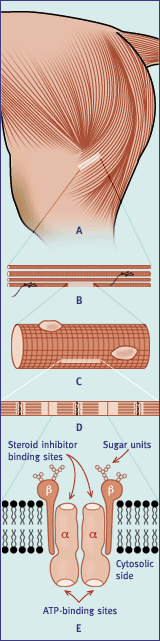
Fig. 1 A - D Schematic representation of the component parts of a
skeletal (striated) muscle. (A) whole muscle. (B) a small part of the muscle
magnified to show the individual muscle cells or fibres. (C) a part of the
muscle fibre magnified. (D) an individual myofibril. A group of myofibrils
together forms a muscle fibre or cell. The Na+,K+ pump is located within
the plasma membrane of these muscle cells. E - Diagrammatic representation
of the subunit structure and orientation of the Na+,K+ pump in the plasma
membrane.
The pump is located within the sarcolemma of skeletal muscle cells (Figure
1, A-D) and is a so-called ab heterodimer, consising
of two a and two b subunit
proteins that have a fixed orientation within the lipid bilayer of the cell
membrane (Figure 1; Part E). It is the larger a
subunit that has the three receptor sites for binding sodium ions on the
portion of the protein protruding to the interior of the cell, the two potassium
ion receptor sites found on the exterior and, adjacent or near to the sodium
ion binding sites, it has the ATPase activity, hence the terms 'pump' and
'enzyme' are often used interchangeably. Like many enzymes,
Na+,K+-ATPase exists in various isoforms, giving rise
to tissue-specific expression and differential regulation of the molecule.
Skeletal muscle expresses a1,
a2, b1 and
b2 isoforms and thus has the possibility of four
ab combinations [41]. The role of the
Na+,K+ pump in the restoration of membrane potential
after exitation [4,5,6], and its confirmational changes during a transport
cycle, in which sodium ions are transported out and potassium ions in to
the cell, are illustrated in Figure 2.
Fig.2 Animation showing a transport
cycle of the Na+,K+ pump. During the cycle, the Na+,K+ pump is first
phosphorylated and subsequently de-phosphorylated. Binding of intracellular
Na+ ions to the pump initiate phosphorylation of the enzyme, while the binding
of extracellular K+ ions trigger the dephosphorylation reaction. After a
complete cycle, three Na+ ions have been extruded from and two K+ ions have
entered into the cell. (Adapted from an illustration by Kjell Lundin, featured
in a special publication to mark the award of the 1997 Nobel Prize in Chemistry
to Professor Jens Christian Skou).
Thus, as a result of repetitive action potentials, exercise induces a loss
of K+ from the muscle cells into the extracellular space, giving
rise to an increase in plasma K+ [4,5,27]. In man, hyperkalemia
occurs during both dynamic and static exercise and is believed to play a
role in the development of muscular fatigue [27,36]. While the long-term
control of plasma K+ concentrations depends ultimately on kidney
function, achieved by increasing the concentration of
Na+,K+ pumps in the cell membrane (for example, by
thyroid hormones or training), it is skeletal muscle that plays the dominant
role in its acute adjustment, by increasing the activity of the
Na+,K+ pump (for example, by adrenaline; Figure 3),
or by increasing the concentration of Na+,K+ pumps
in the cell membrane (for example, by thyroid hormones or training). These
muscles represent the body's largest pool of K+ and
Na+,K+ pumps, and therefore provide an enormous capacity
for rapid Na+,K+ exchange [4,5,6].
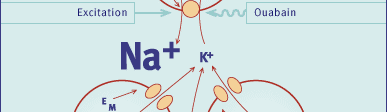
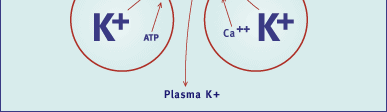
Fig. 3 Diagram of regulatory factors controlling the activity and
the concentration of Na+,K+ pumps in a skeletal muscle cell. Left: factors
eliciting acute stimulation of the Na+,K+ pump (not discussed in text). Right:
factors known to influence the concentration of Na+,K+ pumps by stimulation
or inhibition of their synthesis and insertion into the plasma membrane.
The first two factors (thyroid hormones and training) are discussed in this
review. The lower two muscle cells indicate that K+ leaves the cell during
excitation through ATP or Ca2+-dependent K+ channels.
Abbreviations: IGF-1 - insulin-like growth factor I; CGRP - calcitonin
generelated peptide. (Modified from Reference 4).
An increase in the capacity for active Na+,K+ transport
in skeletal muscle should, however, lead to a 'blunted', or 'dampened', rise
in plasma K+ during exercise (Figure 4), and hence to an improvement
in muscle endurance. Indeed, this does occur in man after sprint training
[17,29]. Furthermore, a correlation exists between maximum O2
uptake, running distance and Na+,K+ pump concentration
in skeletal muscle [14]. It has also been reported that a large increase
in the capacity for active Na+,K+ transport occurs
in the skeletal muscle of patients suffering from hyperthyroidism [4,7,22],
despite the condition being associated normally with increased fatigability
and reduced endurance [13,19].
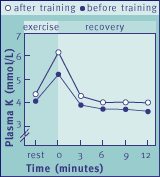
Fig. 4 Changes in plasma K+ concentrations in 4-year old horses (n=3)
during an exercise test performed before and after a 10-day training period.
The rise in plasma K+ during exercise is significantly "blunted" after training.
* P<0.05, after training compared with before training.
Questions
In 1997, the author began investigating the regulation of the
Na+,K+ pump and K+ homeostasis during exercise
in cats, dogs, horses and cattle. Three questions were addressed:
Is thyroid hormone a major determinant in the concentration of
Na+,K+ pumps in skeletal muscle of domestic animal
species, as it is in man?
Does a reduction in Na+,K+ pump concentration lead
to hyperkalemia during exercise? And,
Does training lead to an upregulation in the Na+,K+
pump concentration in skeletal muscle?
Analysis of the concentration of Na+, K+ pumps in
skeletal muscle
Quantitative analysis of membrane bound enzymes, such as
Na+,K+-ATPase, is often performed on plasma membrane
fractions that have been isolated from cell homogenates using differential
centrifugation. However, this procedure requires large amounts of tissue,
making it impractical for both medical and veterinary clinical studies, it
may result in a loss of > 95% of Na+,K+ pumps and
recovery rates between preparations vary enormously [18]. Thus, the development
of techniques to measure the concentration of
Na+,K+ pumps in small samples of intact skeletal muscle
has proved invaluable to physiological studies [30].
Cardiac glycosides, such as digoxin and ouabain, bind specifically to the
outer surface of the Na+,K+ pump, a stoichiometric
process: one molecule of cardiac glycoside binds to one
Na+,K+ pump molecule. The concentration of
Na+,K+ pumps is measured using radioactively-labelled
[3H] ouabain, provided the isozyme of the
Na+,K+ pump in a specific tissue has a high affinity
for the molecule. Analysis of mRNA coding for the
Na+,K+ pump in skeletal muscle has revealed that most
of it does, indeed, code for an isozyme with a high affinity for ouabain
[41].
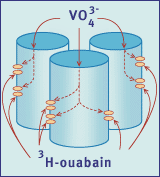
Fig.5 Schematic representation of the vanadate-facilitated binding
of [3H]ouabain to a muscle biopsy. The open ends of the muscle cells allow
vanadate ions access to the phosphorylation site of the Na+,K+ pump, on the
inner surface of the sarcolemma. This binding results in a configuration
of the Na+,K+ pump capable of binding [3H]ouabain to its outer surface (Taken
from Reference 6).
Binding of [3H]ouabain to the Na+,K+ pump
is facilitated by the presence of the phosphate analogue vanadate (Figure
5). Using this anion, a simple and rapid assay has been developed for the
measurement of Na+,K+ pump concentration in muscle
samples weighing as little as 5 mg [30]. The recovery of such small samples
has the considerable advantage of enabling multiple biopsies to be taken
from a tissue and therefore duplicate, triplicate or quadruplicate measurements
may be made.
Thyroid hormones
For about 30 years, it has been known that
Na+,K+-ATPase activity in skeletal muscle and other
tissues increases as a function of thyroid status; hyperthyroidism gives
rise to an increase in pump activity, while hypothyroidism results in its
decreased activity [20]. The increase in Na+,K+ transport
associated with hyperthyroidism was once thought to account for the calorigenic
action of the thyroid hormones, however, only 5-10% of the total heat produced
in skeletal muscle of eu-, hypo- and hyperthyroid animals can be attributed
to active Na+,K+ transport [7].
Na+,K+-ATPase in rat muscle
Thyroid hormones largely determine the concentration of
Na+,K+ pumps in skeletal muscle through a general endocrine
effect [5,7], which is in stark contrast to observations made during training
(see later). In rat skeletal muscle, the
Na+,K+-ATPase concentration is approximately the same
whether it consists predominantly of slow (eg. soleus) or fast (eg.
gastrocnemius) fibres [21]. However, gastrocnemius muscle recovered from
hyperthyroid rats contained five times the concentration of
Na+,K+ pumps compared to equivalent samples recovered
from hypothyroid animals. This difference rose to as much as ten times when
soleus muscle samples were compared. These findings suggest that muscles
show a greater response to an alteration in thyroid status when they consist
predominantly of slow fibres.
Fatigability and Na+,K+ pump capacity
Contrary to expectations, the soleus muscle of hyperthyroid rats shows a
greater susceptibility to fatigue and less endurance [13,19] than its increased
capacity for active Na+,K+-transport suggests [12,19,21].
However, when this phenomenon is considered in relation to the increased
influx of Na+ through specialised channels, it is likely that
muscle endurance is determined by the leak-to-pump ratio of Na+,
not by the Na+,K+ pump concentration alone. Furthermore,
studies in which the time course of the effects of thyroid hormone on
Na+ influx and K+ efflux was compared with that on
Na+,K+-ATPase activity in skeletal muscle, have shown
that the rise in the unidirectional flux of cations preceded the rise in
Na+,K+ pump concentration [12,19]. Thus, increased
permeability of the sarcolemma to cations after thyroid hormone treatment
may be the driving force for the synthesis of
Na+,K+ pumps.
Na+,K+-ATPase in cats
The concentration of Na+,K+ pumps in the skeletal muscles
of hypo- and hyperthyroid dogs and cats has also been determined. Hypothyroidism
is the most frequent thyroid disorder encountered in dogs, while hyperthyroidism
is observed more often in cats [33]. Studies in both these species retrieved
samples from the sternothyroid muscle due to its easy accessibility during
surgical thyroidectomy
Total thyroxine (T4) concentrations were approximately 400% higher,
and Na+,K+ pump concentrations around 75% higher, in
hyperthyroid compared to euthyroid cats (Schaafsma et al, unpublished data).
In both groups of cats, the apparent dissociation constant for ouabain was
of the same order of magnitude as that measured in rats with comparable thyroid
status [21]. An intriguing observation made recently on a cat that was treated
for 10 days with the anti-thyroid drug Strumazol (company, town and country),
showed a high concentration of [3H]ouabain binding sites had been
maintained while the total plasma T4 returned to normal (Schaafsma
et al, unpublished data).
Na+,K+-ATPase and K+ homeostasis in
dogs
Recently, the concentration of Na+,K+ pumps was measured
in the sternothyroid muscle of Beagle dogs, before and after thyroidectomy
[34]. In euthyroid Beagles the Na+,K+ pump concentration
was almost twice that recorded in euthyroid cats, but fell by 40% after
thyroidectomy. The decrease in [3H]ouabain binding capacity was
not due to the Na+,K+ pump's reduced affinity for ouabain.
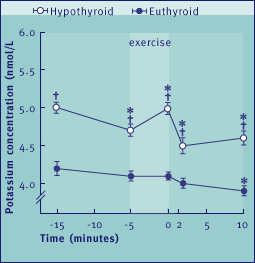
Fig.6 Plasma K+ levels in age-matched euthyroid and hypothyroid Beagle
dogs, before and after exercise. Exercise consisted of a single 5 min run
at a speed of 4.5 km/h. Each point on the graph represents the mean of 6,
or in some cases 7, observations. The bars denote the standard error (SE)
for each mean value;
* P<0.05 or better within groups; † P<0.05 or better between groups.
Total plasma T4 concentrations were about 20 nmol/l in euthyroid
and <2 nmol/l in hypothyroid dogs. The resting plasma K+
concentration was significantly higher in hypothyroid compared to euthyroid
dogs and remained higher throughout the experiment, including the work and
recovery phases of the exercise test (Figure 6). In addition, hypothyroid
dogs showed a significant exercise-induced hyperkalemia. The most likely
explanation for this was a decrease in the muscle's capacity to pump
K+ back into the tissue, since neither muscle damage nor kidney
failure was apparent [34].
Food restriction
Apart from thyroid disorders, thyroid hormone levels may change dramatically
as a result of other diseases or food restriction [10] and may lead to a
change in Na+,K+-ATPase concentration in skeletal muscle.
For example, rats receiving one third to half their normal food supply, for
3 consecutive weeks, revealed a 50% reduction in total plasma triiodothyronine
(T3) in association with a 25% reduction in
Na+,K+-ATPase concentration. This effect proved to
be reversible; after just one week of being fed normal (full) rations, the
rats' plasma T3 and Na+,K+-ATPase concentrations
had returned to normal [4,5]. Similar observations could not be reported
in a group of Shetland ponies subjected to severe, long-term (2.5 years)
food restriction; they showed a reduction in total and free T3,
(30 and 50%, respectively), a proportional loss of body weight, but only
a modest (14%) decrease in Na+,K+-ATPase concentration
in skeletal muscle [38]. This raised the questions whether skeletal muscle
Na+,K+-ATPase isoforms are identical between species
and to what extent thyroid hormone regulates specific isoforms [46].
Training and immobilisation
Depending on its intensity, exercise is accompanied by a rise in plasma
K+ concentrations [27,36], most probably originating from the
working muscles. It is believed that inadequate sarcolemmal
Na+,K+ -ATPase activity and a failure to restore
Na+,K+ gradients across the sarcolemma during excitation
are responsible [4,5,6,27]. Exercise-induced hyperkalemia is reduced by training
in man [17,29], dogs [25], cattle [16] and horses [28] and is most likely
due to an increase in skeletal muscle Na+,K+ pump
concentration; an observation made in many species including rats [23], guinea
pigs [26], man [14,17,29], horses [28,40] and cattle [44]. Alternatively,
the early release of K+ from cells may occur in association with
H+ exchange [45]; in other words, training induces a reduction
in the K+/H+ exchange, if the blunted rise in plasma
K+ witnessed during exercise is to be explained (Figure 4).
Fig. 4 Changes in plasma K+ concentrations in 4-year old horses (n=3)
during an exercise test performed before and after a 10-day training period.
The rise in plasma K+ during exercise is significantly "blunted" after training.
* P<0.05, after training compared with before training.
Training and immobilisation in rodents
Studies in which the concentration of Na+,K+-ATPase
was measured in the skeletal muscles of different species (rats, guinea pigs,
horses, cattle and man) before and after training, showed a relative effect
of between 15 and 50%. It remains to be seen, however, whether this difference
is related to the muscle type, to the relative size of the animals concerned,
or to the duration and type of training.
One study looked at the combined effect of immobility and training on
Na+K+-ATPase concentration in the fast, gastrocnemius,
and slow, soleus, muscles of guinea pigs [26]. Within one to three weeks,
the gastrocnemius muscle Na+,K+-ATPase concentration
had decreased to a maximum of 25% its original value. However, during a fourth
week of immobilisation these levels returned spontaneously to their normal
value. After three weeks of daily running exercise on a treadmill, the
Na+,K+-ATPase concentration increased by 50% in fast
muscle but only by 15% in slow muscle. In rats, six weeks of swimming was
found to induce a comparable (40%) increase in [3H]ouabain binding
site concentration in slow (soleus) and fast (extensor digitorum longus)
muscles [23].
Training studies in man
Studies in man investigating the effects of training on
Na+,K+-ATPase concentration in skeletal muscle, often
involve the collection of biopsies from the easily accessible vastus lateralis
muscle, which consists of mixed types of fibre. Invariably, these studies
use bicycle training as the preferred form of exercise. It not only works
the relevant muscle group sufficiently, but it is also easily standardised
in a laboratory setting. Two simultaneous studies showed an increase of 14%
[17] and 16% [29] in the concentration of [3H]ouabain binding
sites in the vastus lateralis muscle of male subjects, aged 18 to 20 years.
The first of these studies demonstrated this effect after only six, two-hour
daily training sessions [17]. In the second study, in which subjects performed
short bouts of sprint work three times a week, biopsies were not taken until
seven weeks after the start of training [29]. Thus, although the rise in
Na+,K+-ATPase concentration was similar after endurance
and sprint training, a longer period of sprint training was required to attain
this effect. Due to the characteristic mixed fibre type of the vastus lateralis
muscle, the increase in Na+,K+-ATPase concentration
cannot be ascribed to one type of muscle fibre and, because biopsies were
taken only at the end of the seven-week sprint training period, neither can
it be established at what time point changes in
Na+,K+-ATPase concentration occurred first.
Training studies in young and adult horses
Young horses, sprint-trained from birth until five months of age [42], showed
an increase in [3H]ouabain binding capacity in gluteus medius and semitendinosus
muscles of 30% and 20%, respectively [40]. Adult horses also revealed a 36%
rise in Na+,K+-ATPase concentration in the gluteus
medius muscle [20]. In adult horses, this rise was associated with a significant
reduction in the plasma K+ concentration during an exercise test
[28].
Measurements of Na+,K+-ATPase concentrations in the
gluteus medius muscle of young and adult horses affected by periodic hyperkalemic
paralysis have been compared with those of age-matched control horses [31].
It was concluded that the cell membrane events underlying the periodic episodes
of paralysis in hyperkalemic horses could not be attributed to changes in
the Na+,K+ pump in either the
Na+,K+ number or affinity. In addition, the decrease
in Na+,K+-ATPase concentration measured in skeletal
muscle showed an age-dependent decrease. This is true also for rat muscle,
in which the concentration of Na+,K+ pumps rises five-fold
from birth to four weeks of age and then falls due to an increase in the
diameter of mature muscle cells [4,5]. Finally, when
Na+,K+ pump concentrations were compared in gluteus
muscle samples taken from horses of similar age but of different breeds,
including the Quarter horse, Thoroughbred and Dutch warmblood, they were
found to be similar [28,31,40].
Cattle
When endurance-trained Hereford calves were exercised at a maximum sustainable
rate, they showed a rise in peak arterial plasma K+ concentrations
due to an increased maximum work capacity [16]. However, when they were exercised
at a similar work load before and after physical conditioning, the rise was
significantly reduced [16]. Young male and female Mozambican Angoni cattle,
subjected to two hours of draught work every day for two weeks, showed increases
in the concentration of Na+,K+-ATPase in semitendinosus
muscle of 16 and 30%, respectively. When plasma K+ concentrations
were measured regularly during the daily two-hour training periods, the rise
in concentration was lower at the end of the two weeks than it was after
only eight days of training. This difference was not significant however
[44].
Persistence of the training effect
How long does the training-induced rise in
Na+,K+-ATPase concentration persist when intensive
training is discontinued? Rats, subjected to six weeks of swim training,
revealed a large rise in [3H]ouabain binding site concentration,
in both soleus (slow) and extensor digitorum longus (fast) muscles [23],
which was almost completely reversed within three weeks of training being
stopped. However, when a five month training period for young horses was
followed by a six month period of rest, the concentration of
[3H]ouabain binding sites in semitendinosus muscle remained the
same and in gluteus medius muscle was reduced by 10% [39]. Whether this
discrepancy is due to species differences or the type of exercise performed
is difficult to conclude, but the topic warrants further studies.
Is the training effect due to a general or a specific effect?
In trained rats, swimming induced up-regulation of the
Na+,K+-ATPase in all hind limb and spinal muscles,
but not in the diaphragm [23]. This result provides evidence against the
existence of a non-specific endocrine factor, such as thyroid hormone, resposible
for eliciting the training effect on the concentration of
Na+,K+-ATPase [5]. A recent study in young foals has
confirmed this idea by demonstrating that the training-induced rise in
Na+,K+-ATPase was apparent in the gluteus and
semitendinosus muscles of the hind limb, but not in the masseter muscle of
the jaw [39]. Considered together, these observations suggest that the factor
eliciting an up-regulation in the Na+,K+ pump numbers
during training is located in the muscle itself.
Perspectives for future research
In addition to being essential for locomotion, skeletal muscle from some
animals is consumed as meat by man. A muscle's movement and meat quality
are determined by the growth and composition of its composite fibres, as
well as by the maintenance of ion gradients. The physiological and morphological
properties of adult skeletal muscle are the combined result of genetic
predisposition, diet, hormonal influences and the workload that the muscle
has been exposed to.
During development skeletal muscles not only hypertrophy but also adapt to
their required mechanical functions, such as rapid short-lasting movements
(fast muscles) or prolonged actions (slow muscles). With respect to meat
quality, slow muscles have better water holding capacity but lower colour
stability than fast muscles [24].
A muscle's functional adaptation during development is evident through changes
in the cation transport activity [8] and in the myosin heavy chain isoform
expression during the postnatal period [9]. Both parameters are strongly
affected by thyroid hormones and by exercise [5,32,35]. These effects are
not easy to investigate independently since standardising exercise regimes
is difficult and maintaining animals with relatively long growth periods
is costly.
Muscle cell culture
The use of tissue culture techniques to study the adaptive behaviour and
growth of muscle cells has obvious advantages [1]. Foetal myoblasts and adult
muscle satellite cells are readily isolated and grown in vitro (Figure 7).
After an initial phase of proliferation they fuse to form myotubes and then
differentiate to become spontaneously contracting myofibres. During further
growth, the satellite cells divide and their nuclei are added to the fibres,
mimicking the processes of muscle growth and regeneration after injury.
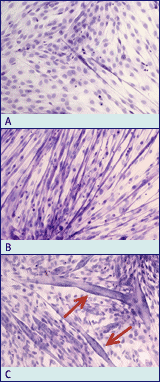
Fig. 7 Myotube formation of skeletal muscle cells grown in culture.
Cell line C2C12 is used to demonstrate this phenomenon. Heamatoxylin-eosin
staining of bouin-fixed muscle cell cultures during several stages of
differentiation. After reaching confluence in medium containing 12% fetal
calf serum, differentiation is induced by growing the cells in medium containing
2% horse serum. Small myotubes are observed after 3 days in culture (A).
By day 7, the number and size of myotubes increases rapidly (B). During the
second week of culture, large multinucleated myotubes are formed (C), and
spontaneous contraction can be observed. Note the peripheral localisation
of the nuclei (arrows). Magnification: x 200 (Photomicrographs provided courtesy
of Dr. Karim Sultan)
A better understanding of the involvement of satellite cells in postnatal
myogenesis and in muscle hypertrophy will be essential to improve the efficiency
of muscle growth in meat producing animals [3]. Muscle cells from pigs [11]
and cattle [3] have been cultured successfully and directed towards
differentiation or proliferation by growth factors [15] and hormones [3].
Electrical and mechanical stimulation have also been applied to the cells
in vitro, as a means of mimicking the application of a 'workload' [43]. Cultured
human skeletal muscle cells have also been analysed for their degree of maturity
by measuring Na+,K+-ATPase activity [2].
Future Research
We are currently developing an in vitro model to test the hypothesis that
exercise, hormones and growth factors together determine the variations found
in growth and fibre composition of skeletal muscle. Two fundamental questions
are under consideration. First, do slow and fast muscle fibres respond
differently to physical and hormonal stimulation? And second, when during
embryonic and post-natal development are the growth and composition of skeletal
muscle fibres most affected by these stimuli?
Concluding remarks
The concentration of Na+,K+ pumps in the skeletal muscle
of cats, dogs, horses and cattle is regulated by mechanisms similar to those
described in rodents and man. We already know that hyperthyroidism and physical
training increase the number of Na+,K+ pumps in skeletal
muscle and that hypothyroidism and immobility reduce their number. We know,
too, that the rise in Na+,K+ pump concentration after
training is associated with a blunted rise in plasma K+ during
exercise. However, the question remains whether the mechanism responsible
for the up-regulation of the Na+,K+ pump during
hyperthyroidism is the same as that during training.
References
1. Allen, R.A. and Rankin, L.A. (1990) Regulation of satellite
cells during skeletal muscle growth and development.
Proc. Soc. exp. Biol. Med. 194, 81-86.
2. Benders, A.A.G.M., van Kuppevelt, T.H.M.S.M., Oosterhof,
A., Wevers, R.A., Veerkamp, J.H. (1992) Adenosine triphosphatase during
maturation of cultured human skeletal muscle cells and in adult human muscle.
Biochim. Biophys. Acta 1112, 89-98.
3. Cassar-Malek, I., Langlois, N., Picard, B. and Geay, Y.
(1999) Regulation of bovine satellite cell proliferation and differentiation
by insulin and triiodothyronine.
Domest. Anim. Endocrinol. 17, 373-388.
4. Clausen, T. (1996) Long and short-term regulation of the
Na+,K+ pump in skeletal muscle. News Physiol. Sci.
11, 24-30.
5. Clausen, T. and Everts, M.E. (1989) Regulation of the
Na+,K+ pump in skeletal muscle.
Kidney Int. 35, 1-13.
6. Clausen, T. and Nielsen, O.B. (1999) The
Na+,K+-pump in muscle and CNS - physiology and
pathophysiology. NS 2, 4-8.
7. Clausen, T., van Hardeveld, C. and Everts, M.E. (1991)
Significance of cation transport in control of energy metabolism and
thermogenesis.
Physiol. Rev. 71, 733-774.
8. Dauncey, M.J. and Harrison, A.P. (1996) Developmental regulation
of cation pumps in skeletal and cardiac muscle.
Acta Physiol. Scand. 156, 313-323.
9. Dingboom, E.G., Dijkstra, G., Enzerink, E., van Oudheusden,
H.C. and Weijs, W.A. (1999) Postnatal muscle fibre composition of the gluteus
medius muscle of Dutch warmblood foals; maturation and the influence of exercise.
Equine vet. J. Supplement 31, 95-100.
10. Docter, R., Krenning, E.P. de Jong, M. and Hennemann,
G. (1993) The sick euthyroid syndrome: Changes in thyroid hormone serum
parameters and hormone metabolism. Clin. Endocrinol. 39, 499-518.
11. Doumit,M.E. and Merkel, R.A. (1992) Conditions for isolation
and culture of porcine myogenic satellite cells.
Tissue and Cell 24, 253-262.
12. Everts, M.E. and Clausen, T. (1988) Effects of thyroid
hormone on Na+,K+-transport in resting and stimulated
rat skeletal muscle.
Am. J. Physiol. 255, E604-E612.
13. Everts, M.E., Simonides, W.S., Leijendekker, W.J. and
van Hardeveld, C. (1987) Fatigability and recovery of rat soleus muscle in
hyperthyroidism.
Metabolism 36, 444-450.
14. Evertsen, F., Medbø, J.I., Jebens, E. and Nicolaysen,
K. (1997) Hard training for 5 months increases
Na+,K+ pump concentration in skeletal muscle of
cross-country skiers.
Am. J. Physiol. 272, R1417-R1424.
15. Fligger, J.M., Malven, P.V., Doumit, M.E., Merkel, R.A.
and Grant, A.L. (1998) Increases in insulin-like growth factor binding protein-2
accompany decreases in proliferation and differentiation when porcine muscle
satellite cells undergo multiple passages.
J. Anim. Sci. 76, 2086-2093.
16. Fosha-Dolezal, S.R. and Fedde, M.R. (1988) Serum potassium
during exercise in Hereford calves: influence of physical conditioning.
J. appl. Physiol. 65, 1360-1366.
17. Green, H.J., Chin, E.R., Ball-Burnett, M. and Ranney,
D. (1993) Increases in human skeletal muscle
Na+,K+-ATPase concentration with short-term training.
Am. J. Physiol. 264, C1538-C1541.
18. Hansen, O. and Clausen, T. (1988) Quantitative determination
of the Na+,K+-ATPase and other sarcolemmal components
in muscle cells.
Am. J. Physiol. 254, C1-C7.
19. Harrison, A.P. and Clausen, T. (1998) Thyroid hormone-induced
upregulation of Na+ channels and Na+,K+
pumps: implications for contractility.
Am. J. Physiol. 274, R864-R867.
21. Kjeldsen, K., Everts, M.E. and Clausen, T. (1986a) The
effects of thyroid hormones on 3H-ouabain binding site concentration,
Na+,K+ contents and 86Rb-efflux in rat skeletal
muscle.
Pflugers Arch. 406, 529-535.
22. Kjeldsen, K., Norgaard, A., Gotzsche, C.O., Thomassen,
A., Clausen, T. (1984) Effect of thyroid function on number of
Na+-K+ pumps in human skeletal muscle.
Lancet II, 8-10.
23. Kjeldsen, K., Richter, E.A., Galbo, H., Lortie, G. and
Clausen, T. (1986b) Training increases the concentration of 3H-ouabain binding
sites in rat skeletal muscle.
Biochim. Biophys. Acta 860, 708-712.
24. Klont, R.E., Brocks, L. and Eikelenboom, G. (1998) Muscle
fibre type and meat quality. Meat Science 49 Supplement 1, S219-S229.
25. Knochel, J.P., Blachley, J.D., Johnson, J.H. and Carter,
N.W. (1985) Muscle cell electrical hyperpolarization and reduced exercise
hyperkalemia in physically conditioned dogs.
J. clin. Invest. 75, 740-745.
26. Leivseth, G., Clausen, T., Everts, M.E. and Bjorda, E.
(1992) Effects of reduced joint mobility and training on
Na+,K+-ATPase and Ca2+-ATPase in skeletal
muscle.
Muscle and Nerve 15, 843-849.
27. Lindinger, M.I. and Sjøgaard, G. (1991) Potassium
regulation during exercise and recovery.
Sports Med. 11, 382-401.
28. McCutcheon, L.J., Geor, R.J. and Shen, H. (1999) Skeletal
muscle Na+,K+-ATPase and K+ homeostasis
during exercise: effects of short-term training.
Equine vet. J. Supplement 30, 303-310.
29. McKenna, M.J., Schmidt, T.A., Hargreaves, M., Cameron,
L., Skinner, S.L. and Kjeldsen, K. (1993) Sprint training increases human
skeletal muscle Na+,K+-ATPase concentration and improves
K+ regulation.
J. appl. Physiol. 75, 173-180.
30. Nørgaard, A., Kjeldsen, K. and Clausen, T. A. (1984)
Method for the determination of the total number of 3H-ouabain binding sites
in biopsies of human skeletal muscle.
Scand. J. clin. lab. Invest. 44, 509-518.
31. Pickar, J.G., Spier, S.J. Harrold, D. and Carlsen, R.C.
(1993) [3H]Ouabain binding in skeletal muscle from horses with
hyperkalemic periodic paralysis.
Am. J. vet. Res. 54, 783-787.
32. Putman, C.T., Dusterhoft, D. and Pette, D. (1998) Changes
in satellite cell content and myosin isoforms in low-frequency-stimulated
fast muscle of hypothyroid rats.
J.
appl. Physiol. 86, 40-51.
33. Rijnberk, A. (1996) The thyroids. In: Clinical endocrinology
of dogs and cats: an illustrated text. Ed. A. Rijnberk. Kluwer Academic
Publishers, Amsterdam, pp. 35-59.
34. Schaafsma, I.A., van Emst, M.G., Kooistra, H.S., Verkleij,
C.B., Peeters, M.E., Rijnberk, A. and Everts, M.E. (2000) Exercise induced
hyperkalemia in hypothyroid dogs. Abstracts of the World Congress of
WSAVA/FECAVA, Amsterdam, 2000.
35. Simonides, W.S. and van Hardeveld, C. (1989) The postnatal
development of sarcoplasmic reticulum Ca2+-transport activity
in skeletal muscle of the rat is critically dependent on thyroid hormone.
Endocrinology 124, 1145-1153.
36. Sjøgaard, G. (1990) Exercise-induced muscle fatigue:
the significance of potassium. Acta Physiol. Scand. 140, Suppl. 593, 1-63.
37. Skou, J.C. (1957) The influence of some cations on an
adenosine triphosphatase from peripheral nerves. Biochim. Biophys. Acta 23,
394-401.
38. Suwannachot, P., Verkleij, C.B., Kocsis, S., Enzerink,
E. and Everts, M.E. (2000) Prolonged food restriction and mild exercise in
Shetland ponies: effects on weight gain, thyroid hormone concentrations,
and muscle Na+,K+-ATPase.
J. Endocrinol. In press.
39. Suwannachot, P., Verkleij, C.B., Kocsis, S., van Weeren,
P.R. and Everts, M.E. (2000) Specificity and reversibility of the training
effects on the concentration of Na+,K+-ATPase in foal
muscle. Equine vet. J. In press.
40. Suwannachot, P., Verkleij, C.B., Weijs, W.A., van Weeren,
P.R. and Everts, M.E. (1999) Effects of training on the concentration of
Na+,K+-ATPase in foal muscle. Equine vet. J. Suppl
31, 101-105.
42. van Weeren, P.R. and Barneveld, A. (1999) The influence
of exercise during the first months of life on the development of the equine
musculoskeletal system with special attention to osteochondrosis: Experimental
set-up and scope of the study.
Equine vet. J. Suppl 31, 4-8.
43. Vandenburgh, H.H. (1992) Mechanical forces and their second
messengers in stimulating cell growth in vitro.
Am. J. Physiol. 262, R350-R355.
44. Veeneklaas, R.J., Verkleij, C.B., van Schie, B., Harun,
M.A.S. and Everts, M.E. (2000) Na+,K+-ATPase in skeletal
muscle of draught animals in Mozambique: effect of race, gender, age and
training. Submitted for publication.
45. Wasserman, K., Stringer, W.W. Casaburi, R. and Zhang,
Y-Y. (1997) Mechanism of the exercise hyperkalemia: an alternate hypothesis.
J. appl. Physiol. 83, 631-643.
46. Yalcin, Y., Carman, D., Shao, Y., Ismail-Beigi, F., Klein,
I. and Ojamaa, K. (1999) Regulation of
Na+/K+-ATPase gene expression by thyroid hormone and
hyperkalemia in the heart.
Thyroid 9, 53-59.